
Oxygen is the final electron acceptor in the body. It binds with the electrons at the end of the electron transport chain in respiration. Therefore the transport of oxygen across the body is a crucial procedure as oxygen must be able to travel to all the respiring cells in the body to collect the electrons at the end of respiration. Thats why there are specialised proteins which are able to load and unload oxygen at will. Without these very crucial proteins life wouldn’t be able to survive.
We are often told of haemoglobin and its role in oxygen transport. Haemoglobin isn’t the only oxygen transport protein in the body however. Another protein found in muscles specifically does a similar role to haemoglobin and it’s name is myoglobin. The functional differences between the two proteins occur because of their structure which we will look at below.

Haemoglobin is the transporter protein found in the blood, the red blood cells specifically. It’s role is to load oxygen from the lungs and unload it at respiring tissue and load carbon dioxide from respiring tissue and offload it in the lungs to be expelled out of the body. This whole mechanism is supported by the structure of haemoglobin. It’s a globular protein that is tetrameric – consists of 4 polypeptide chains (2 alpha chains and 2 beta chains) and within each of these chains exist a prosthetic (non-protein) heme group. The existence of the heme group in haemoglobin allows it to bind to the oxygen molecules at will. There being 4 of them means that haemoglobin can bind to a maximum of 4 oxygen molecules. When it is fully saturated it is usually referred to as oxyhaemoglobin and when it isn’t saturated with oxygen it can be referred to as deoxyhaemoglobin. With the deoxyhemoglobin variant, there are two states in which it can exist: Low affinity T state or the high affinity R state. The binding of oxygen to the Iron (II) ion in the heme group promotes the stabilisation of the R state leading to a conformational change causing it to go into the R state.
Myoglobin, unlike haemoglobin, consists of a single polypeptide chain, hence it is known as a monomeric protein. It has a heme group (characteristic of proteins which transport oxygen) and is a globular structure protein composed of 8 separate right handed alpha helices connected via non-helical regions. As aforementioned, myoglobin contains a heme group which in itself contains the Iron (II) ion to which the oxygen binds to. When bonded to oxygen it’s referred to as oxymyoglobin and when unbounded to oxygen it’s referred to as oxymyoglobin.
Now we have briefly discussed the structures of these two proteins we will now look at the functions they have in the body. To get a general understanding it’s important to note that for both haemoglobin and myoglobin, the affinity for oxygen can change depending on a few environmental factors.
Myoglobin is mainly used as a storage molecule for oxygen. It is found within muscle tissue, and is most important during oxygen deprivation. This is why it has an extremely high affinity for oxygen hence it binds to oxygen very tightly. In these anaerobic situations, where there is oxygen deprivation, the oxygen released from myoglobin can allow the muscle cells to carry on generating energy to contract. Due to this it is very easily saturated at low partial pressures of oxygen. When plotting a graph of saturation against partial pressure of oxygen (also known as a oxygen dissociation curve), we can identify the curve for myoglobin as being hyperbolic.

Haemoglobin, in comparison, has a lower affinity to oxygen than myoglobin. This allows the release of oxygen into tissue, to aid with the function of haemoglobin. The oxygen dissociation curve for haemoglobin tends to be of a sigmoidal shape. This is due to cooperative binding – as haemoglobin gets more saturated, it is easier for oxygen molecules to bind to haemoglobin, hence affinity to oxygen increases as the haemoglobin gets more saturated.

The sigmoidal shape of the curve shows that oxygen can be efficiently transported around the body, allowing the red blood cell to carry out its function. At the lungs, hemoglobin will have a high affinity for oxygen as the partial pressure is high. This means that near the lungs it will be close to 100% saturated. However, as the red blood cell moves down the blood and transports oxygen to peripheral tissues, partial pressure of oxygen decreases, hence the affinity of oxygen also decreases. Hemoglobin becomes less saturated.
P50 is the partial pressure at which 50% of the hemoglobin is saturated. The p50 value for hemoglobin is a lot lower than that for myoglobin. This is significant as the myoglobin will continue to hold up its oxygen until oxygen deprivation (low pO2) occurs in the muscles. In comparison, hemoglobin will unload its oxygen molecules at a higher partial pressure of oxygen as it delivers the oxygen more readily to respiring tissues. This also allows hemoglobin to give up its oxygen to myoglobin – as myoglobin stores the oxygen molecules.
There are a few factors that alter the regulation of oxygen in hemoglobin. These include carbon dioxide concentration and pH, temperature, binding of BPG and type of hemoglobin.
- Carbon Dioxide concentration and pH
Carbon Dioxide is produced by aerobic respiration. This causes the concentration of carbon dioxide to increase in blood, hence more carbon dioxide molecules bind to the haemoglobin in red blood cells. Concentration is linked to H+ ions by the catalytic reaction which occurs with carbon anhydrase:
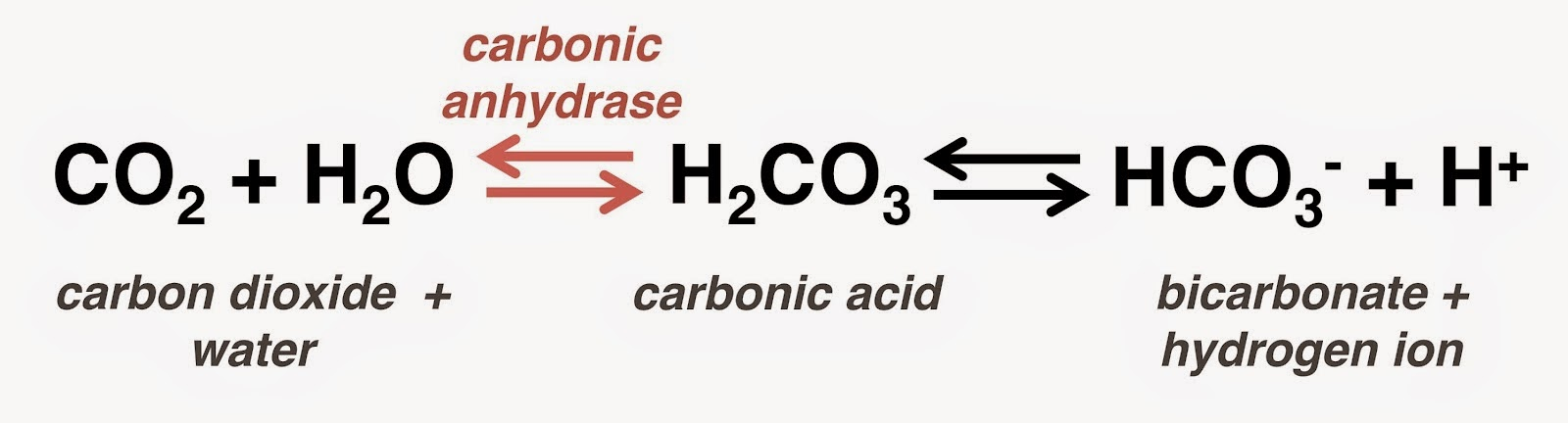
From this equation, we can conclude that as the carbon dioxide concentration increases, the H+ ion concentration also increases. This causes a decrease in pH.
Both carbon dioxide and H+ ions bind to hemoglobin away from the oxygen binding site and cause the hemoglobin to enter a relaxed T state. Carbon dioxide molecules bind to the terminal amino acid subunits forming carbaminohemoglobin, whereas H+ binds to amino acid chains and makes up the actual globin subunits. This causes a decrease in affinity with oxygen, forcing oxygen to unload. Let’s take the example of muscle cells. When the continuously work, they release a lot of CO2, hence a lot of H+ ions. This causes a reduction of pH, leading to the unloading of oxygen to the muscle cells which need the oxygen to respire at that point.
When CO2 concentration is increased the saturation curve will shift to the right to compensate for the decrease in affinity.
- Temperature
Temperature affects the strength of the bond between the oxygen and the Fe2+ in the centre of the subunits. The Fe2+ ion acts as a sort of anchor of oxygen to hemoglobin. As the temperature increases, the bond is weakened causing the hemoglobin to favour the T state. This reduces affinity to oxygen, causing the unloading of oxygen and a decreased saturation.
In this case the saturation curve will shift to the right to compensate for the decrease in affinity.
- Binding of 2,3-Biphosphoglycerate (BPG)
BPG is an intermediate product of glycolysis. The abundance of BPG increases at hypotoxic conditions – an example is this is at high altitudes where there is a lack of oxygen. Just like CO2 and H+ ions, BPG does not bind to the oxygen sites of hemoglobin. It similarly favours the T state, therefore decreasing affinity of Hb and in turn decreasing saturation of oxygen.
The dissociation curve will also shift to the right to compensate for te decrease in affinity.
- Type of Hemoglobin
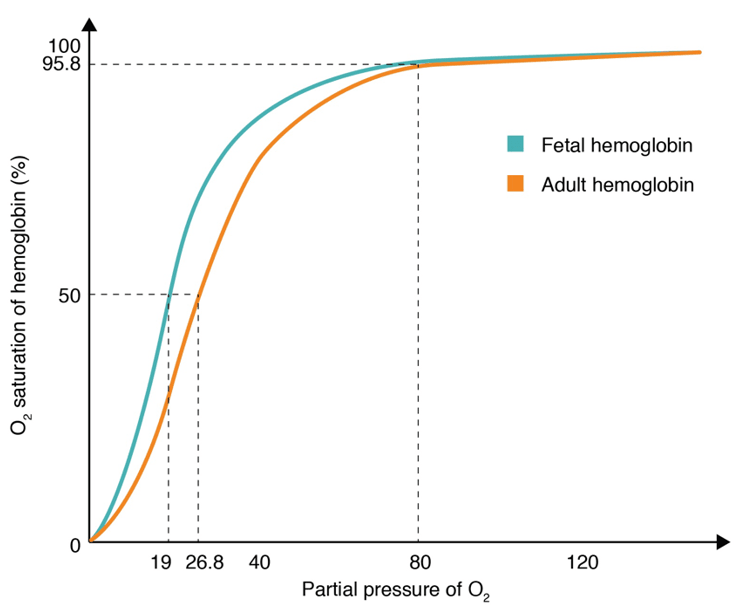
Prenatal hemoglobin is actually different to the hemoglobin found in adults. This is due to a different regulation of Hb genes. Normally in adults, the Hb is composed of four subunits: 2 alpha and 2 beta subunits. In fetal hemoglobin the beta subunits are actually replaced with 2 gamma subunits. This causes a change in the properties the hemoglobin possesses.
Fetal hemoglobin tends to have a much higher affinity than adult hemoglobin. This allows efficient transfer of oxygen from the blood of the mother to the blood of the child. The oxygen is transferred from maternal oxyhemoglobin from via the placenta. The fetal hemoglobin prevents the binding of 2,3-BPG to red blood cells, due to the 2 gamma subunits which replaced the beta subunits. The oxygen dissociation curve is shifted to the left, when compared to adult hemoglobin.
- Carbon Monoxide
Carbon monoxide binds to the myoglobin and haemoglobin at the same site which the oxygen binds to, only it binds more tightly. Because of this tight binding, carbon monoxide isn’t released easily by the proteins and therefore the proteins can’t continue with their role of oxygen transport as the carbon monoxide displaces the oxygen on the protein. After one carbon monoxide molecule binds to one subunit of haemoglobin, it increases the affinity of unaffected subunits for oxygen and therefore three oxygens bind to the haemoglobin with carbon monoxide. However, as the affinity of oxygen in this haemoglobin is so high, it is unable to release these oxygens thus this haemoglobin is rendered useless. As more and more carbon monoxide bind to the haemoglobin, there are less haemoglobin present to release oxygen to respiring cells. Therefore cells can be starved of oxygen. Common symptoms include: Dizziness, tiredness and unconsciousness. It becomes fatal when there are >50COHb molecules are present in the blood.

Oxygen transport is a crucial procedure. It’s a necessity to support life itself. Therefore understanding the processes and proteins involved helps us to paint a picture of how our body works: the procedures that are involved and the tools which are used to execute said procedures. Having this knowledge enables us to try and look for solutions whenever things go wrong and so is fundamental knowledge to aspiring medics and scientists alike.

Leave a comment